LED Lights Promote Growth and Flavonoid Accumulation of Anoectochilus roxburghii and Are Linked to the Enhanced Expression of Several Related Genes

 ,
,  and
and
Abstract
:
1. Introduction
2. Results
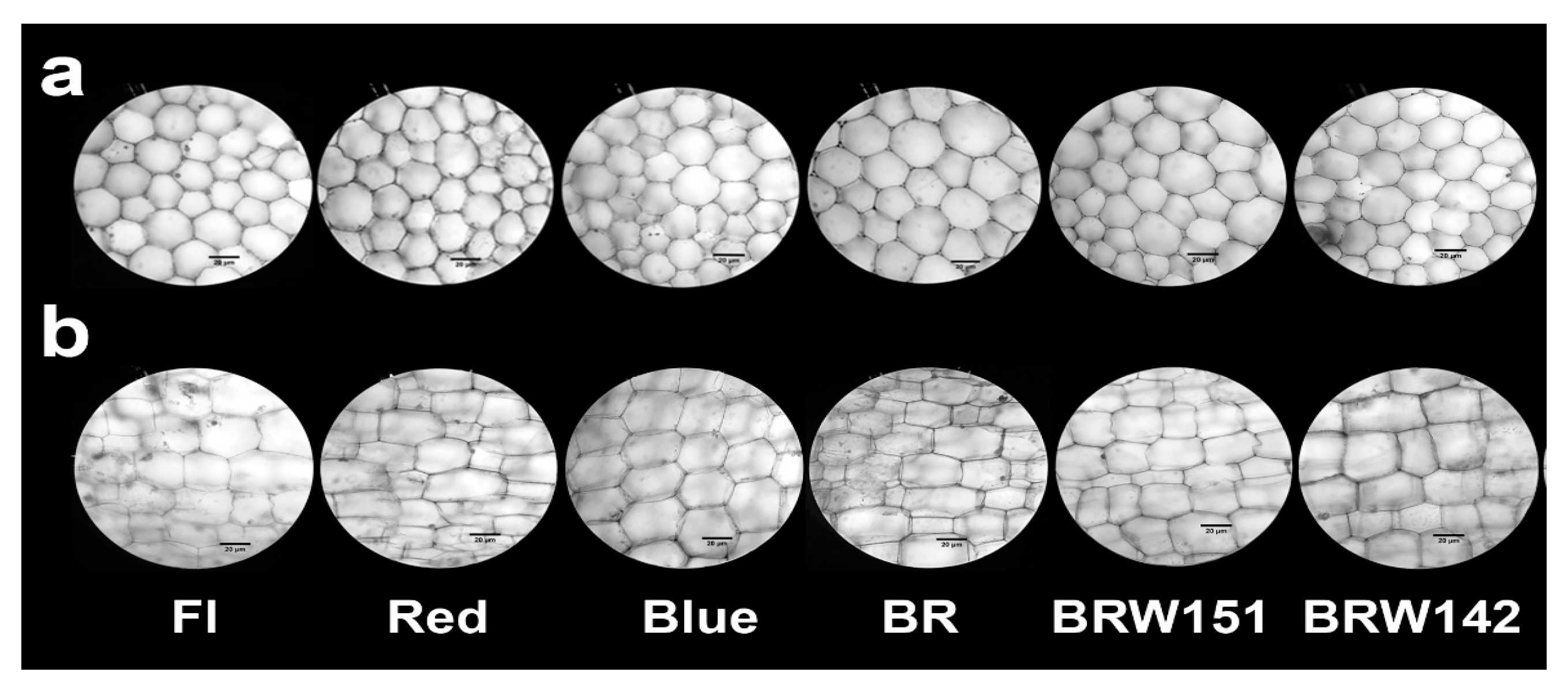
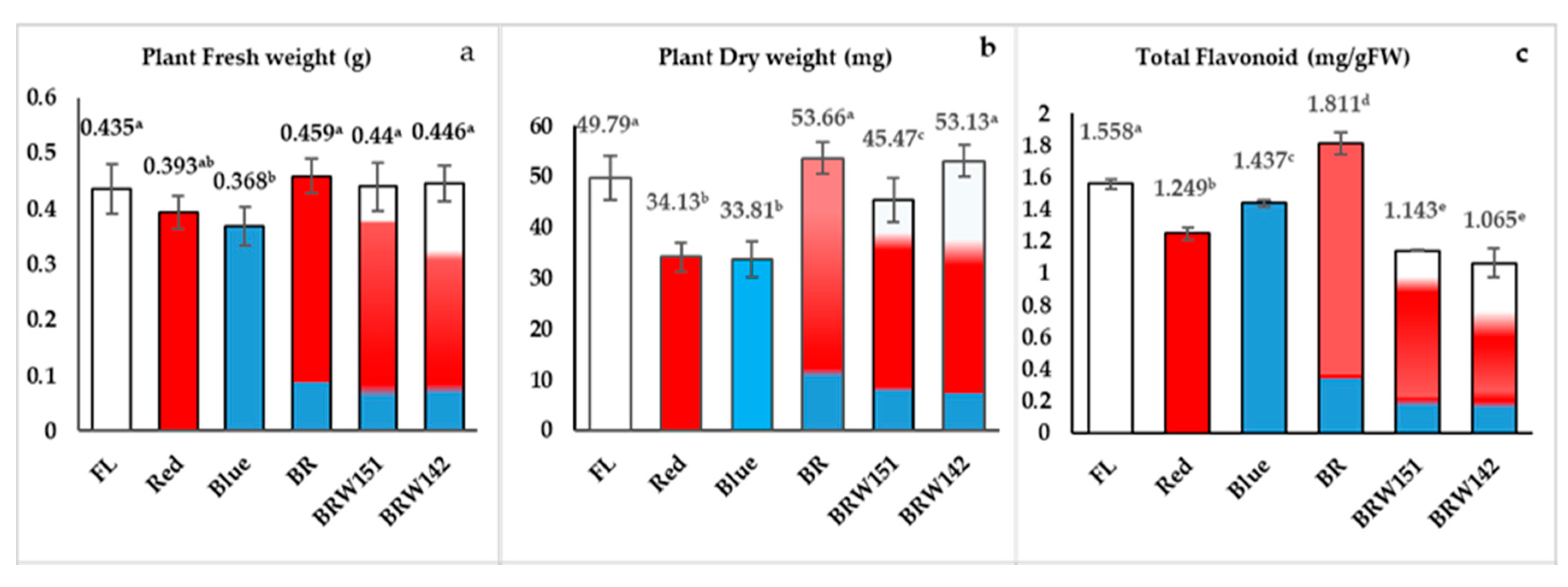
2.1. Effects of LED Light Treatments on Growth and Morphological Characteristics
2.2. Effects of LED Lights on Total Flavonoids Accumulation
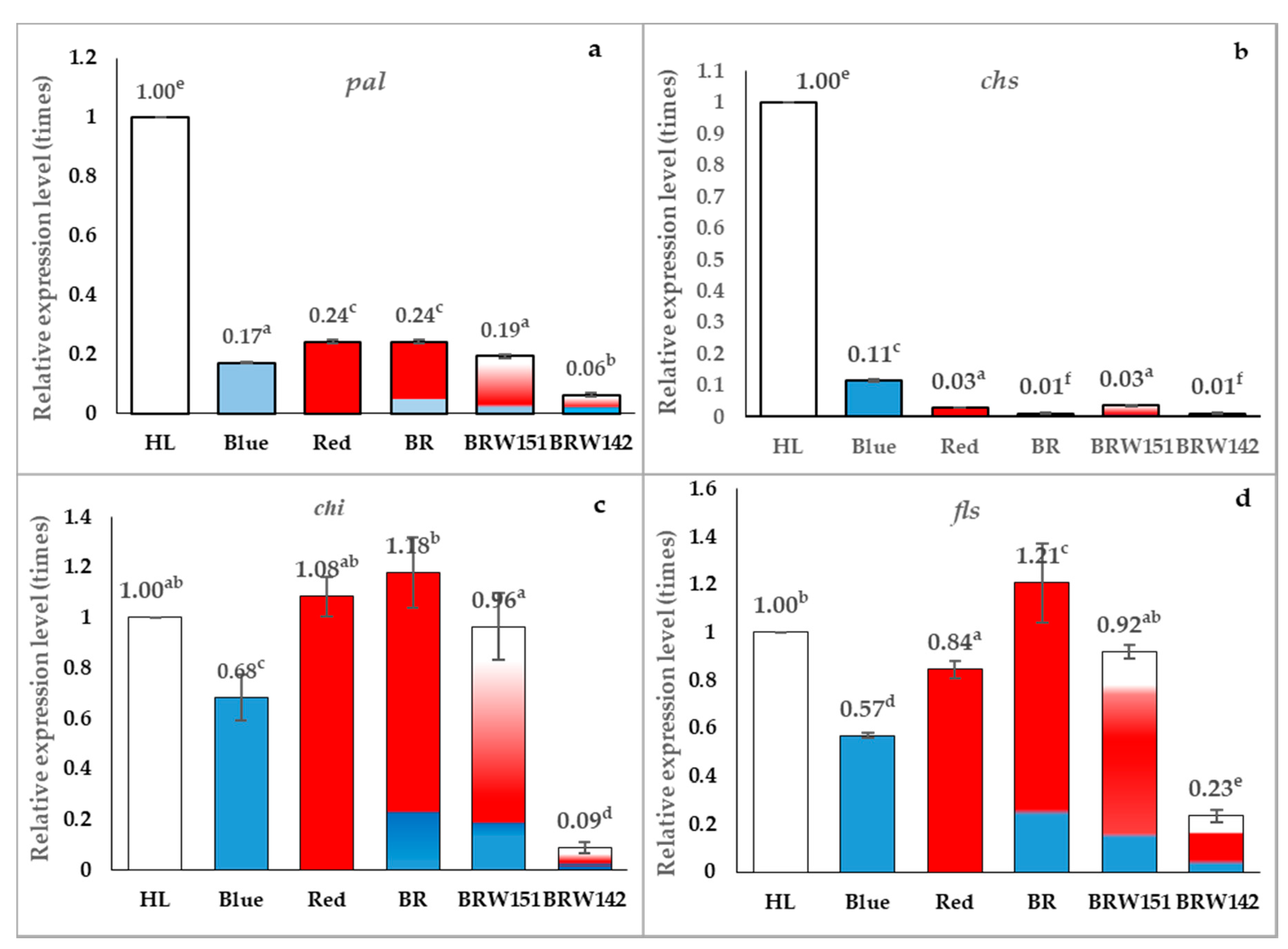
2.3. Effects of LED Light on Expression Level of pal, chs, chi, and fls Encoding Genes
3. Discussion
4. Materials and Methods
4.1. Plants Materials and Growth Conditions
4.2. Morphological and Biomass Parameter Analyses
4.3. Stem Anatomical Characteristics
4.4. Determination of Total Flavonoids Contents
4.5. Expression Analysis of Several Flavonoid Biosynthesis-Related Genes
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, Y.; Cai, J.; Ruan, H.; Pi, H.; Wu, J. Antihyperglycemic activity of kinsenoside, a high yielding constituent from Anoectochilus roxburghii in streptozotocin diabetic rats. J. Ethnopharmacol. 2007, 14, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.C.; Yu, J.; Zhang, X.H.; Cheng, M.Z.; Yang, L.W.; Xu, J.Y. Antihyperglycemic and antioxidant activity of water extract from Anoectochilus roxburghii in experimental diabetes. Exp. Toxicol. Pathol. 2013, 65, 485. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Wang, H.; Guo, H.; Zhou, A.; Huang, Y.; Sun, Y.; Li, M. Effects of shade treatments on photosynthetic characteristics, chloroplast ultrastructure, and physiology of Anoectochilus roxburghii. PLoS ONE 2014, 9, e85996. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Shao, Q.; Xu, M.; Li, S.; Wu, M.; Tan, X.; Su, L. Effects of light quality on morphology, enzyme activities, and bioactive compound contents in Anoectochilus roxburghii. Front. Plant Sci. 2017, 8, 857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Y.; Chen, Y.F.; Wang, L.H.; Hsu, K.Y.; Chin, Y.T.; Yu-Chen, S.H.Y.; Wang, S.H.; Chen, Y.R.; Shih, Y.J.; Liu, L.F.; et al. Inhibitory effect of Anoectochilus formosanus extract on hyperglycemia-Related PD-L1 expression and cancer proliferation. Front. Pharmacol. 2018, 9, 87. [Google Scholar] [CrossRef]
- National Compendium of Chinese Herbal Medicine, Editorial Board. National Compendium of Chinese Herbal Medicine; People’s Medical Publishing House Beijing: Beijing, China, 1978. [Google Scholar]
- Lin, W.C.; Shih, C.C.; Wu, Y.W.; Chen, I.J.; Chang, H.L.; Hsu, C.T. Ninety-day oral toxicity study of crude aqueous extracts of Anoectochilus formosanus Hayata in rats. J. Chin. Med. 2000, 11, 1–22. [Google Scholar]
- Xu, W.J.; Chen, Y.; Huang, Z.Q.; Yang, S.Y.; Chen, J.Y. A preliminary study on the treatment of compound herb a anoectochili capsule on diabetes II. Subtrop. Plantence 2000, 29, 47–49. [Google Scholar]
- Liu, Z.F.; Li, Q. Clinical Observation on 30 cases of chronic hepatitis B treated by compound Jinlianlian oral liquid combined with entecavir. Fujian J. TCM 2008, 39, 3–4. [Google Scholar]
- Xiao, Z.W. Treatment of 60 cases of infantile cough variant asthma with Anoectochilus roxburghii mixture. Chin. Arch. Tradit. Chin. Med. 2004, 22, 20–25. [Google Scholar]
- Chen, X.X.; Xia, X.N.; Zhang, D.F.; Huang, M.; Xiao, Z.G. Treatment of 36 cases of hyperuricemia in elderly patients with Anoectochilus roxburghii capsule. Mil. Med. J. Southeast China 2010, 12, 331–332. [Google Scholar]
- Li, Q.; Zhou, W.; Liu, L.; Zheng, L.; Guan, S.C. Clinical observation on the treatment of hand-foot-mouth disease with Anoectochilus roxburghii spray. Fujian J. TCM 2012, 43, 9–10. [Google Scholar]
- Xiao, W. Treatment of 49 cases of infantile Tourette syndrome with Anoectochilus roxburghii oral liquid. J. Northwest Pharmacol. 2005, 20, 81–82. [Google Scholar]
- Yan, Y.B. Clinical observation on treatment of 60 cases of Hp infection with omeprazole and Anoectochilus roxburghii. Fujian J. TCM 2008, 39, 11–12. [Google Scholar]
- Tang, F.; Zhang, X.Q.; Xu, J.T.; Cai, J.Y.; Zhang, J.W.; Zhang, Y.H. Screening on hypoglycemic effective part of Anoectochilus Roxburghii. Chin. Trad. Herb. Drugs 2011, 42, 340–342. [Google Scholar]
- Hsiao, H.B.; Wu, J.B.; Lin, H.; Lin, W.C. Kinsenoside isolated from Anoectochilus Formosanus suppresses LPS-stimulated inflammatory reactions in macrophages and endotoxin shock in mice. Shock 2011, 35, 184–190. [Google Scholar] [CrossRef]
- Huang, L.F.; Lu, R.Y.; Su, Z.M.; Fan, S.T.; Yu, X. Effect of herba Anoectochili extracts on acutely and chronically damaged livers induced by CCl_4 in mice. Pharm. J. Chin. Peoples Lib. Army 2007, 23, 278–281. [Google Scholar]
- Ye, S.; Shao, Q.; Zhang, A. Anoectochilus Roxburghii: A review of its phytochemistry, pharmacology, and clinical applications. J. Ethnopharmacol. 2017, 209, 184–202. [Google Scholar] [CrossRef]
- Shang wen, L.; Yanyan, W.; Shishi, H.; Xuenong, Z.; Yang, Z.; Yihong, X.; Changxing, Q.; Weiguang, S.; Yongbo, X.; Jianping, W.; et al. The water extract of Anoectochilus formosanus Protects mice from concanavalin A-induced hepatitis. Int. J. Clin. Exp. Med. 2018, 11, 9210–9219. [Google Scholar]
- Do, T.G.; Ha, V.H.; Chu, H.H.; Pham, B.N. Investigation of some chemical characteristics and antioxidant effects of flavonoid compounds extracted from the species of Anoectochilus in Vietnam. VNU J. Sci. Nat. Sci. Technol. 2017, 33, 104–113. [Google Scholar]
- Park, C.; Kim, N.; Park, J.; Lee, S.; Lee, J.W.; Park, S. Effects of light-Emitting diodes on the accumulation of glucosinolates and phenolic compounds in sprouting canola (Brassica Napus L.). Foods 2019, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Abidi, F.; Girault, T.; Douillet, O.; Guillemain, G.; Sintes, G.; Laffaire, M.; Ben-Ahmed, H.; Smiti, S.; Huché-Thélier, L.; Leduc, N. Blue light effects on rose photosynthesis and photomorphogenesis. Plant Biol. 2013, 15, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Kozai, T.; Niu, G.; Takagaki, M. Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Nhut, D.T.; Hong, L.T.A.; Watanabe, H.; Goi, M.; Tanaka, M. Growth of banana plantlets cultured in vitro under red and blue light-emitting diodes (LED) irradiation source. Acta Hortic. 2002, 575, 117–123. [Google Scholar] [CrossRef]
- Zhang, L.C.; Ma, G.; Yamawaki, K.; Ikoma, Y.; Matsumoto, H.; Yoshioka, T.; Ohta, S.; Kato, M. Effect of blue LED light intensity on carotenoid accumulation in citrus juice sacs. J. Plant Physiol. 2015, 188, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Y.; Jiao, X.L.; Chang, T.T.; Guo, S.R.; Xu, Z.G. Photosynthesis and leaf development of cherry tomato seedlings under different LED-based blue and red photon flux ratios. Photosynthetica 2018, 56, 1212–1217. [Google Scholar] [CrossRef]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Mohidul, H.; Tufail, B.; Ritesh, G.; Sun, K.L.; Hanhong, B. An Overview of LEDs’ Effects on the Production of Bioactive Compounds and Crop Quality. Molecules 2017, 22, 1420. [Google Scholar]
- Lekkham, P.; Srilaong, V.; Pongprasert, N.; Kondo, S. Anthocyanin concentration and antioxidant activity in light-emitting diode (LED)-treated apples in a greenhouse environmental control system. Fruits 2016, 71, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Azad, M.O.K.; Kim, W.W.; Park, C.H.; Cho, D.H. Effect of Artificial LED light and far infrared irradiation on phenolic compound, isoflavones and antioxidant capacity in soybean (Glycine max L.) Sprout. Foods 2018, 7, 174. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Su, M.; Li, H.; Zeng, B.; Chang, Q.; Lai, Z. Effects of supplemental lighting with different light qualities on growth and secondary metabolite content of Anoectochilus roxburghii. PeerJ 2018, 6, e5274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoma, F.; Somborn-Schulz, A.; Schlehuber, D.; Keuter, V.; Deerberg, G. Effects of Light on Secondary Metabolites in Selected Leafy Greens: A Review. Front. Plant Sci. 2020, 11, 497. [Google Scholar] [CrossRef]
- Ma, Z.H.; Li, W.F.; Mao, J.; Li, W.; Zuo, C.W.; Zhao, X.; Dawuda, M.M.; Shi, X.Y.; Chen, B.H. Synthesis of light-inducible and light-independent anthocyanins regulated by specific genes in grape ‘Marselan’ (V. Vinifera L.). PeerJ 2019, 7, e6521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.H.; Zheng, X.T.; Sun, B.Y.; Peng, C.L.; Chow, W.S. Over-expression of the CHS gene enhances resistance of Arabidopsis leaves to high Light. Environ. Exp. Bot. 2018, 154, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Ghasemzadeh, A.; Jaafar, H.Z.E.; Rahmat, A. Synthesis of phenolic and flavonoids in Ginger (Zingiber officinale Roscoe) and their effects on photosynthesis rate. Int. J. Mol. Sci. 2010, 11, 4539–4555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Li, S.; Wu, X.; Zhang, J.; He, A.; Zhao, G.; Yang, X. Effect of light intensity on leaf photosynthetic characteristics and accumulation of flavonoids in Lithocarpus litseifolius (Hance). Chun. (Fagaceae). Open J. For. 2016, 6, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Karimi, E.; Jaafar, H.Z.E.; Ghasemzadeh, A.; Ibrahim, M.H. Light intensity effects on production and antioxidant activity of flavonoids and phenolic compounds in leaves, stems and roots of three varieties of Labisia pumila benth. Aust. J. Crop Sci. 2013, 7, 1016–1023. [Google Scholar]
- Yuan, M.; Jia, X.; Ding, C.; Zeng, H.; Du, L.; Yuan, S.; Zhang, Z.; Wu, O.; Hu, C.; Liu, J. Effects of fluorescence light on phenolic compounds and antioxidant activities of soybeans (Glycine max L. Merrill) during germination. Food Sci. Biotechnol. 2015, 24, 1859–1865. [Google Scholar] [CrossRef]
- Lv, T.; Teng, R.; Shao, Q.; Wang, H.; Zhang, W.; Li, M.; Zhang, L. DNA barcodes for the identification of Anoectochilus roxburghii and its adulterants. Planta 2015, 242, 1167–1174. [Google Scholar] [CrossRef]
- Goto, E. Plant production in a closed plant factory with artificial lighting. Acta Hortic. 2012, 956, 37–49. [Google Scholar] [CrossRef]
- Xu, Y. Nature and source of light for plant factory. In Plant Factory Using Artificial Light Adapting to Environmental Discruption and Clues to Agricultural Innovation; Masakazu, A., Fukuda, H., Wada, T., Eds.; Elsevier Publishing: Cambridge, MA, USA, 2019; pp. 47–69. [Google Scholar]
- Heo, J.; Lee, C.; Chakrabarty, D.; Paek, K. Growth responses of marigold and salvia bedding plants as affected by monochromic or mixture radiation provided by a Light-Emitting Diode (LED). Plant Growth Regul. 2002, 38, 225–230. [Google Scholar] [CrossRef]
- Su, N.; Wu, Q.; Shen, Z.; Xia, K.; Cui, J. Effects of light quality on the chloroplastic ultrastructure and photosynthetic characteristics of cucumber seedlings. Plant Growth Regul. 2014, 73, 227–235. [Google Scholar] [CrossRef]
- Macedo, A.F.; Leal-Costa, M.V.; Tavares, E.S.; Lage, C.L.S.; Esquibel, M.A. The effect of light quality on leaf production and development of in vitro cultured plants of Alternanthera brasiliana Kuntze. Environ. Exp. Bot. 2011, 70, 43–50. [Google Scholar] [CrossRef]
- Simlat, M.; Lêzak, P.; Mos, M.; Warcho, M.; Skrzypek, E.; Ptak, A. The effect of light quality on seed germination, seedling growth and selected biochemical properties of Stevia rebaudiana Bertoni. Sci. Hortic. 2016, 211, 295–304. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Halimah, N.; Ko, C.H.; Jeong, B.R. Blue LED light enhances growth, phytochemical contents, and antioxidant enzyme activities of Rehmannia glutinosa cultured in vitro. Hortic. Environ. Biotechnol. 2015, 56, 105–113. [Google Scholar] [CrossRef]
- Ryu, J.H.; Seo, K.S.; Choi, G.L.; Rha, E.S.; Lee, S.C.; Choi, S.K.; Kang, S.Y.; Bae, C.H. Effects of LED light illumination on germination, growth and anthocyanin content of Dandelion (Taraxacum officinale). Korean J. Plant Res. 2012, 25, 731–738. [Google Scholar] [CrossRef] [Green Version]
- Doi, K.; Izawa, T.; Fuse, T.; Yamanouchi, U.; Kubo, T.; Shimatani, Z.; Yano, M.; Yoshimura, A. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes Dev. 2004, 18, 926–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H. Phytochromes and light signal perception by plants? An emerging synthesis. Nature 2000, 407, 585–591. [Google Scholar] [CrossRef]
- Devlin, P. “Photoconttrol of Flowering”. In Light and Plant Development; Whitelam, G.C., Halliday, K.J., Eds.; Blackwell: Oxford, UK, 2007; pp. 185–210. [Google Scholar]
- Jericó, J.B.B.; Eduardo, M.E.; Jos Humberto, C.V.; Victorino, M.R. Effect of LED Light quality on in vitro shoot proliferation and growth of Vanilla (Vanilla Planifolia Andrews). Afr. J. Biotechnol. 2016, 15, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Hu, L.; Jiang, D.; Xi, W. Effect of post-harvest LED and UV light irradiation on the accumulation of flavonoids and limonoids in the segments of Newhall Navel Oranges (Citrus Sinensis Osbeck). Molecules 2019, 24, 1755. [Google Scholar] [CrossRef] [Green Version]
- Gould, K.; Davies, K.M.; Winefield, C. Anthocyanins: Biosyntheis, Function and Application; Winefield, C., Davies, K., Gould, K., Eds.; Springer: New York, NY, USA, 2009. [Google Scholar]
- Guo, H.; Wang, Y.; Wang, L.; Hu, P.; Wang, Y.; Jia, Y.; Zhang, C.; Zhang, Y.; Zhang, Y.; Wang, C.; et al. Expression of the MYB transcription factor gene BplMYB46 affects abiotic stress tolerance and secondary cell wall deposition in Betula platyphylla. Plant Biotechnol. J. 2017, 15, 107–121. [Google Scholar] [CrossRef]
- Besseau, S.; Hoffmann, L.; Geoffroy, P.; Lapierre, C.; Pollet, B.; Legrand, M. Flavonoid accumulation in Arabidopsis repressed in lignin synthesis affects auxin transport and plant growth. Plant Cell 2007, 19, 148–162. [Google Scholar] [CrossRef] [Green Version]
- Thain, S.C.; Murtas, G.; Lynn, J.R.; McGrath, R.B.; Millar, A.J. The circadian clock that controls gene expression in Arabidopsis is tissue specific. Plant Physiol. 2002, 130, 102–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Shen, H.; Xu, H.; Tang, X.; Tang, M.; Ju, Z.; Yi, Y. Chalcone isomerase a key enzyme for anthocyanin biosynthesis in Ophiorrhiza Japonica. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Millar, A. Input signals to the plant circadian clock. J. Exp. Bot. 2003, 55, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.H.; Lee, H. MYB-related transcription factors function as regulators of the circadian clock and anthocyanin biosynthesis in Arabidopsis. Plant Signal. Behav. 2016, 11, e1139278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Dong, L.; Jiang, Z.; Yang, X.; Chen, Z.; Wu, Y.; Xu, M. Comprehensive transcriptome analysis and flavonoid profiling of Ginkgo leaves reveals flavonoid content alterations in day–night cycles (Min, X.J., Ed.). PLoS ONE 2018, 13, e0193897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, H. Cultivation technology for vegetable and herb production. In Plant Factory Using Artificial Light Adapting to Environmental Discruption and Clues to Agricultural Innovation; Masakazu, A., Fukuda, H., Wada, T., Eds.; Elsevier Publishing: Cambridge, MA, USA, 2019; pp. 15–23. [Google Scholar]
- Easlon, H.M.; Bloom, A.J. Easy leaf area: Automated digital image analysis for rapid and accurate measurement of leaf Area. Appl. Plant Sci. 2014, 2, 1400033. [Google Scholar] [CrossRef]
- Marinova, D.; Ribarova, F.; Atanassova, M. Total phenolics and total flavonoids in Bulgarian fruits and vegetables. J. Uni. Chem. Technol. Metall. 2005, 40, 255–260. [Google Scholar]
- Sultana, B.; Anwar, F.; Ashraf, M. Effect of extraction solvent/technique on the antioxidant activity of selected medicinal plant extracts. Molecules 2009, 14, 2167–2180. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.; Rahmat, A.; Wahab, P.; Halim, M. Effect of different light intensities on total phenolics and flavonoids synthesis and antioxidant activities in young ginger varieties (Zingiber Officinale Roscoe). Int. J. Mol. Sci. 2010, 11, 3885–3897. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, J.C.; Qu, J.T.; He, G.; Yu, H.Q.; Li, W.C.; Fu, F.L. Expression response of chalcone synthase gene to inducing conditions and its effect on flavonoids accumulation in two medicinal species of Anoectochilus. Sci. Rep. 2019, 9, 20171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhao, M.; Song, C.; Luo, A.; Bai, J.; Guo, S. Characterization of reference genes for quantitative real-time PCR analysis in various tissues of Anoectochilus Roxburghii. Mol. Biol. Rep. 2012, 39, 5905–5912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.; Chen, X.; Meng, Z.; Guo, S. Combined metabolome and transcriptome analyses reveal the effects of Mycorrhizal fungus ceratobasidium Sp. AR2 on the flavonoid accumulation in Anoectochilus roxburghii during different growth stages. Int. J. Mol. Sci. 2020, 21, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, Y.; Guo, S. Effects of the Mycorrhizal fungus ceratobasidium Sp. AR2 on growth and flavonoid accumulation in Anoectochilus roxburghii. PeerJ 2020, 8, e8346. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Conditions | FL | Red | Blue | BR | BRW151 | BRW142 |
|---|---|---|---|---|---|---|
| Average of plant height (cm) * | 6.58 a ± 0.38 | 7.36 c ± 0.92 | 7.23 c ± 0.5 | 7.18 bc ± 0.27 | 6.65 a ± 0.42 | 6.99 abc ± 0.78 |
| Average of stem diameter (mm) * | 16.7 cd ± 2.57 | 11.4 a ± 1.39 | 15.6 c ± 2.77 | 17.6 d ± 1.99 | 16.9 bc ± 2.02 | 16.6 cd ± 1.8 |
| Number of leaves per plant * | 5.13 a ± 0.35 | 5.06 a ± 0.59 | 5.53 bc ± 0.51 | 5.87 c ± 0.63 | 5.33 ab ± 0.48 | 5.8 c ± 0.56 |
| Cell width of 2nd node sections (µm) ** | 28.58 c ± 3.2 | 25.37 a ± 4.4 | 36.87 b ± 3.1 | 30.31 d± 3.7 | 33.11 e ± 3.6 | 27.64 bc ± 3.8 |
| Cell length of 2nd node sections (µm) ** | 35.94 bc ±5.6 | 44.33 e ± 5.5 | 31.23 a ± 3.9 | 44.2 e ± 7.4 | 31.98 ab ± 5.1 | 37.82 cd ± 8.5 |
| Leaf area (cm2) * | 3.36 c ± 0.34 | 1.99 a ± 0.32 | 3.02 bc ± 0.43 | 4.67 d ± 0.89 | 3.61 c ± 0.36 | 3.37 c ± 0.7 |
| Number of roots per plant * | 1.73 a ± 0.45 | 2.00 ab ±1.04 | 2.06 bc ± 0.25 | 1.8 ab ± 0.41 | 1.93 ab ± 0.25 | 2.26 cd ± 0.45 |
| Average of roots length (cm) * | 2.94 bc ±0.43 | 2.47 a ± 0.54 | 3.24 c ± 0.47 | 2.8 ab ± 0.45 | 2.94 bc ± 0.49 | 3.23 c ± 0.34 |
| Dry matter percentages (%) * | 11.45 | 8.68 | 9.18 | 11.69 | 10.33 | 11.91 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gam, D.T.; Khoi, P.H.; Ngoc, P.B.; Linh, L.K.; Hung, N.K.; Anh, P.T.L.; Thu, N.T.; Hien, N.T.T.; Khanh, T.D.; Ha, C.H. LED Lights Promote Growth and Flavonoid Accumulation of Anoectochilus roxburghii and Are Linked to the Enhanced Expression of Several Related Genes. Plants 2020, 9, 1344. https://doi.org/10.3390/plants9101344
Gam DT, Khoi PH, Ngoc PB, Linh LK, Hung NK, Anh PTL, Thu NT, Hien NTT, Khanh TD, Ha CH. LED Lights Promote Growth and Flavonoid Accumulation of Anoectochilus roxburghii and Are Linked to the Enhanced Expression of Several Related Genes. Plants. 2020; 9(10):1344. https://doi.org/10.3390/plants9101344
Chicago/Turabian StyleGam, Do Thi, Phan Hong Khoi, Pham Bich Ngoc, Ly Khanh Linh, Nguyen Khac Hung, Phan Thi Lan Anh, Nguyen Thi Thu, Nguyen Thi Thu Hien, Tran Dang Khanh, and Chu Hoang Ha. 2020. "LED Lights Promote Growth and Flavonoid Accumulation of Anoectochilus roxburghii and Are Linked to the Enhanced Expression of Several Related Genes" Plants 9, no. 10: 1344. https://doi.org/10.3390/plants9101344